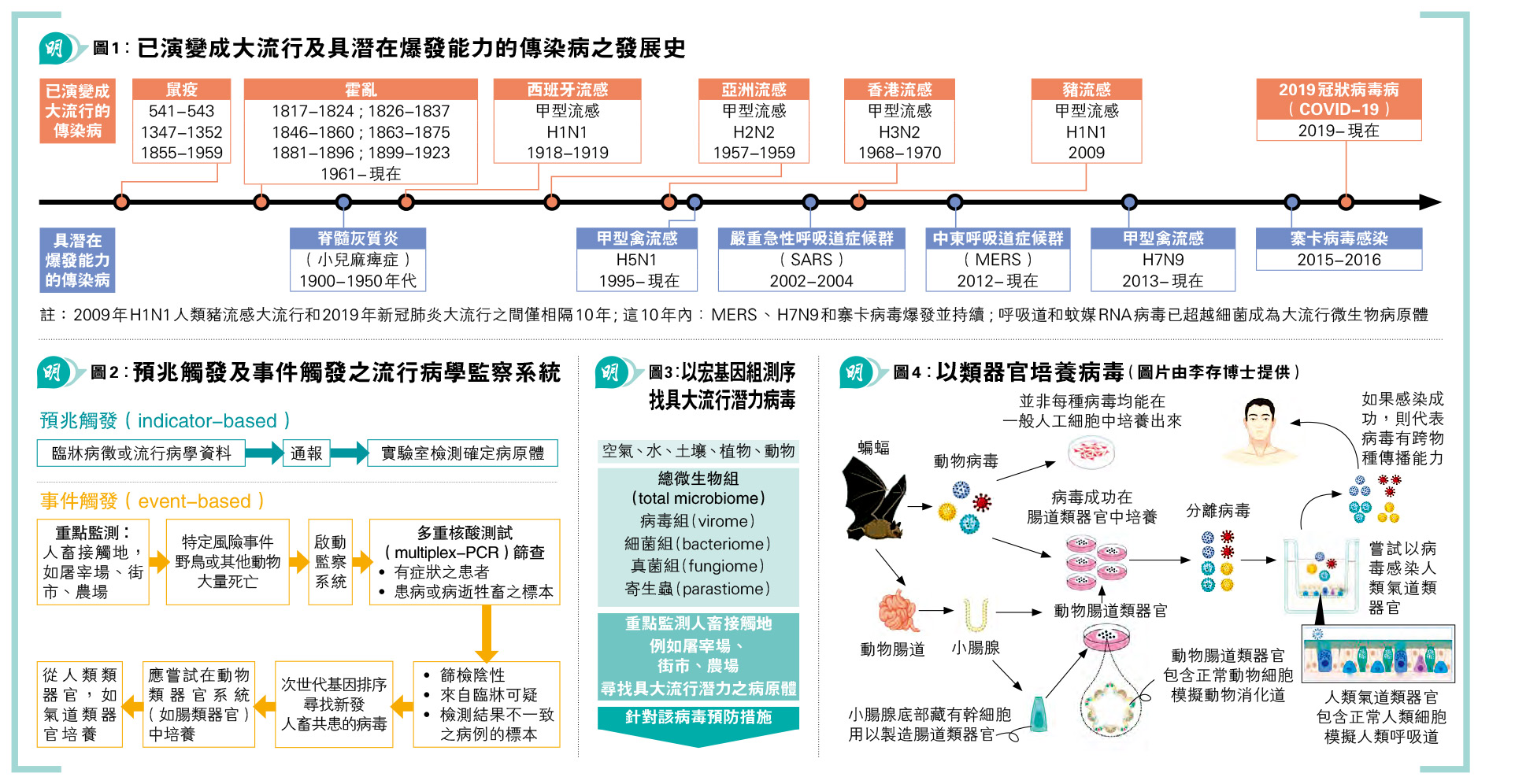
新冠完結,全球復常。但新冠病毒並非最後一場大流行,新發傳染病定必陸續有來。為未來的大流行作準備,必須有高質科研、足夠的基礎建設、迅速應變計劃及措施,完善公營醫療機構、公共衛生及流行病學監察系統,時刻與國際組織保持聯繫溝通等,缺一不可。香港大學將聯同各地學者(包括內地、美國、澳洲及新加坡等),成立大流行科研聯盟(Pandemic Research Alliance),藉此共同尋找具潛力病源,追蹤新發傳染病,應對未來大流行。行政長官於上星期向立法會發表《2023年施政報告》,認為香港必須加強抗擊傳染病能力,但如何用有限度資源去達到監測、預警和防控卻是一門學問。本文綜合本團隊早前於醫學期刊發表之醫學評論文章,從而討論流行病學監察系統(註1)。
頻繁的基因突變會導致抗原漂移(antigenic drift)及分段基因重組(gene reassortment),即俗稱的病毒基因洗牌。此變異使抗原出現轉移,令病毒能重新感染具免疫力的群體,甚至適應不同的宿主細胞環境,從而出現跨物種傳播(cross species transmission),例如2009年豬流感H1N1病毒,以及H5N1和H7N9等禽流感病毒(註7至9)。除人類冠狀病毒OC43和HKU1可能起源於囓齒類動物外,其餘人類冠狀病毒,包括229E、NL63、MERS-CoV、SARS-CoV-1和SARS-CoV-2均被認為是從蝙蝠透過中間宿主傳播到人類(註10)。
註1:Chiu KH, Sridhar S, Yuen KY. Preparation for the next pandemic: challenges in strengthening surveillance. Emerg Microbes Infect. 2023 Dec;12(2):2240441.
註2:Chan JF, Yuan S, Kok KH, et al. A familial cluster of pneumonia associated with the 2019 novel coronavirus indicating person-to-person transmission: a study of a family cluster. Lancet. 2020 Feb 15;395(10223):514-523.
註3:Cheng VC, Lau SK, Woo PC, et al. Severe acute respiratory syndrome coronavirus as an agent of emerging and reemerging infection. Clin Microbiol Rev. 2007 Oct;20(4):660-94.
註4:Chan JF, Lau SK, To KK, et al. Middle East respiratory syndrome coronavirus: another zoonotic betacoronavirus causing SARS-like disease. Clin Microbiol Rev. 2015 Apr;28(2):465-522.
註5:To KK, Sridhar S, Chiu KH, et al. Lessons learned 1 year after SARS-CoV-2 emergence leading to COVID-19 pandemic. Emerg Microbes Infect. 2021 Dec;10(1):507-535.
註6:Zhang X, Chen LL, Ip JD, et al. Omicron sublineage recombinant XBB evades neutralising antibodies in recipients of BNT162b2 or CoronaVac vaccines. Lancet Microbe. 2022 Dec 6.
註7:Cheng VC, To KK, Tse H, et al. Two years after pandemic influenza A/2009/H1N1: what have we learned? Clin Microbiol Rev. 2012 Apr;25(2):223-63.
註8:Peiris JS, Yu WC, Leung CW, et al. Re-emergence of fatal human influenza A subtype H5N1 disease. Lancet. 2004 Feb 21;363(9409):617-9.
註10:Lau SK, Woo PC, Li KS, et al. Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A. J Virol. 2015 Mar;89(6):3076-92.
註11:Woo PC, Lau SK, Tsang CC, et al. Coronavirus HKU15 in respiratory tract of pigs and first discovery of coronavirus quasispecies in 5'-untranslated region. Emerg Microbes Infect. 2017 Jun 21;6(6):e53.
註12:Woo PC, Lau SK, Lam CS, et al. Discovery of seven novel Mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J Virol. 2012 Apr;86(7):3995-4008.
註13:Yuen KY, Chan PK, Peiris M, et al. Clinical features and rapid viral diagnosis of human disease associated with avian influenza A H5N1 virus. Lancet. 1998 Feb 14;351(9101):467-71.
註14:Chen Y, Liang W, Yang S, et al. Human infections with the emerging avian influenza A H7N9 virus from wet market poultry: clinical analysis and characterisation of viral genome. Lancet. 2013 Jun 1;381(9881):1916-25.
註15:Guan Y, Zheng BJ, He YQ, et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science. 2003 Oct 10;302(5643):276-8.
註16:Li C, Huang J, Yu Y, et al. Human airway and nasal organoids reveal escalating replicative fitness of SARS-CoV-2 emerging variants. Proc Natl Acad Sci U S A. 2023 Apr 25;120(17):e2300376120.













 發表於
發表於 












